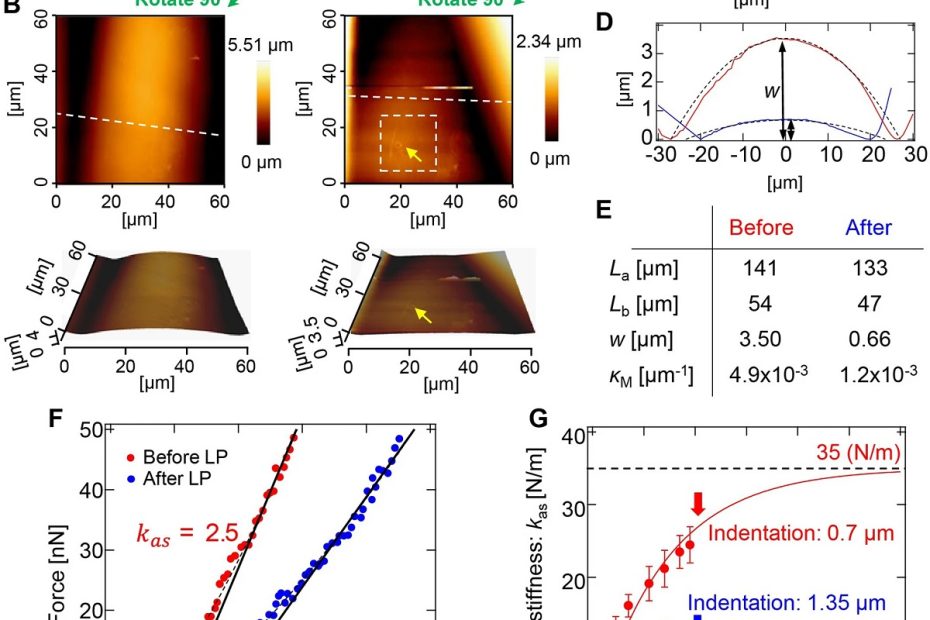
Dynamics of Compartment-Specific Proteomic Landscapes of Hepatotoxic and Cholestatic Models of Liver Fibrosis
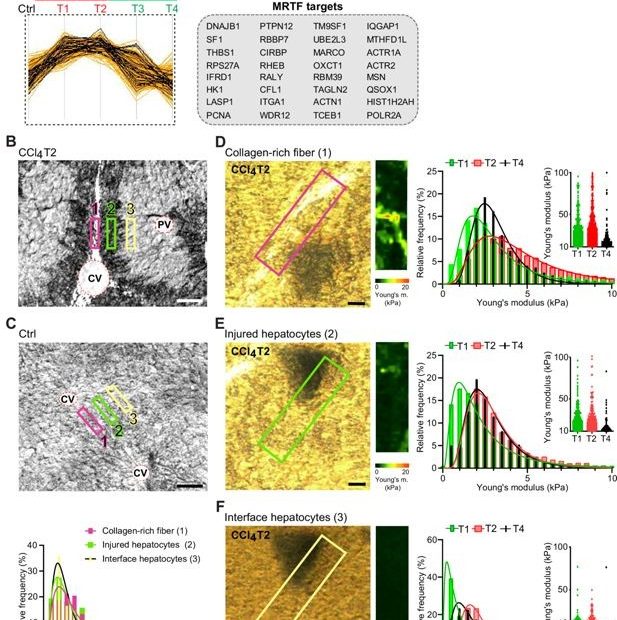
Liver fibrosis is characterized by excessive accumulation and remodeling of extracellular matrix (ECM) components, leading to altered tissue mechanics and progressive impairment of liver function.… Read More »Dynamics of Compartment-Specific Proteomic Landscapes of Hepatotoxic and Cholestatic Models of Liver Fibrosis