Decoding Morphology in Organic Solar Cells with NANOSENSORS™ PPP-NCHAu AFM Probes
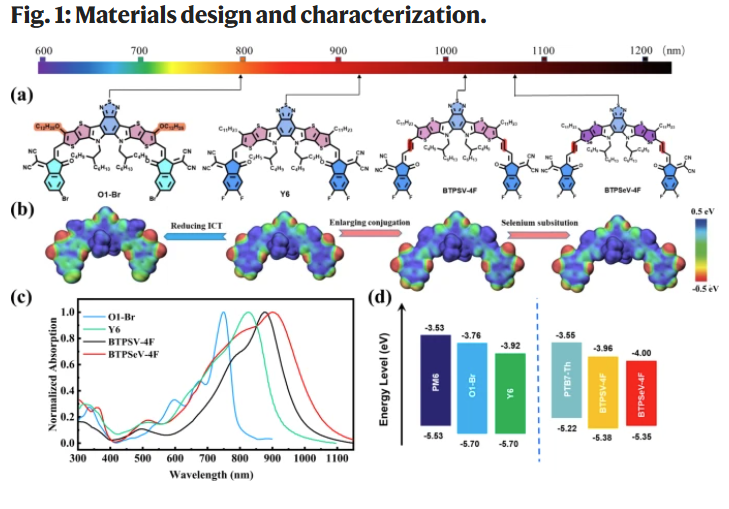
Published in Nature Communications (March 2023), the article by Zhenrong Jia et al. investigates a novel ultra-narrow bandgap non-fullerene acceptor, BTPSeV-4F, which effectively suppresses triplet… Read More »Decoding Morphology in Organic Solar Cells with NANOSENSORS™ PPP-NCHAu AFM Probes