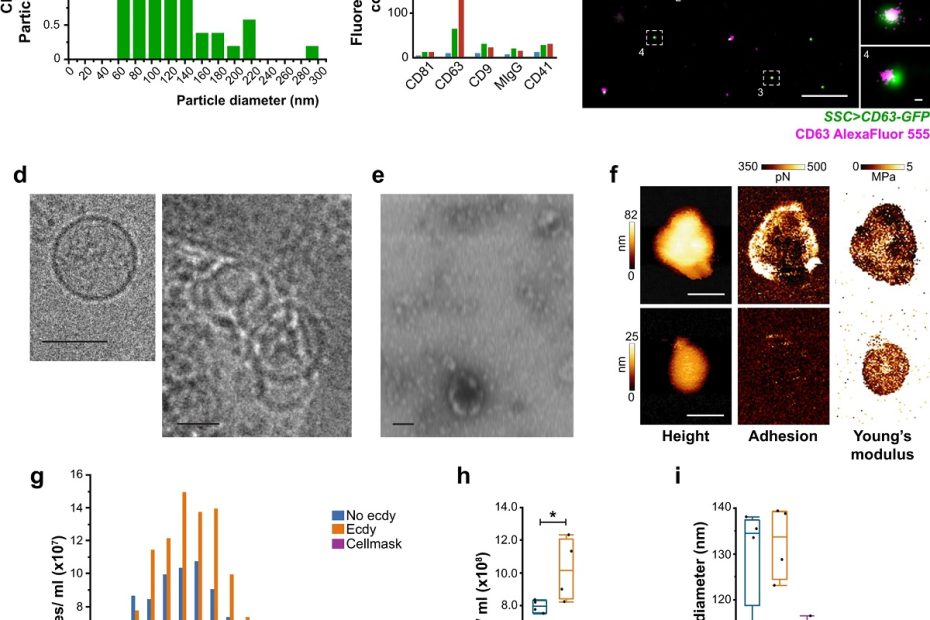
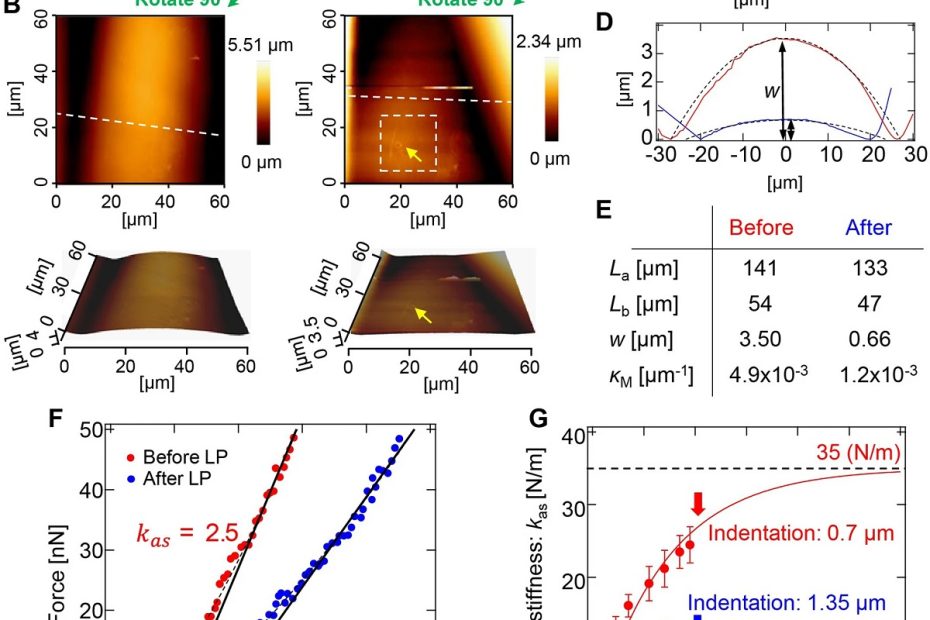
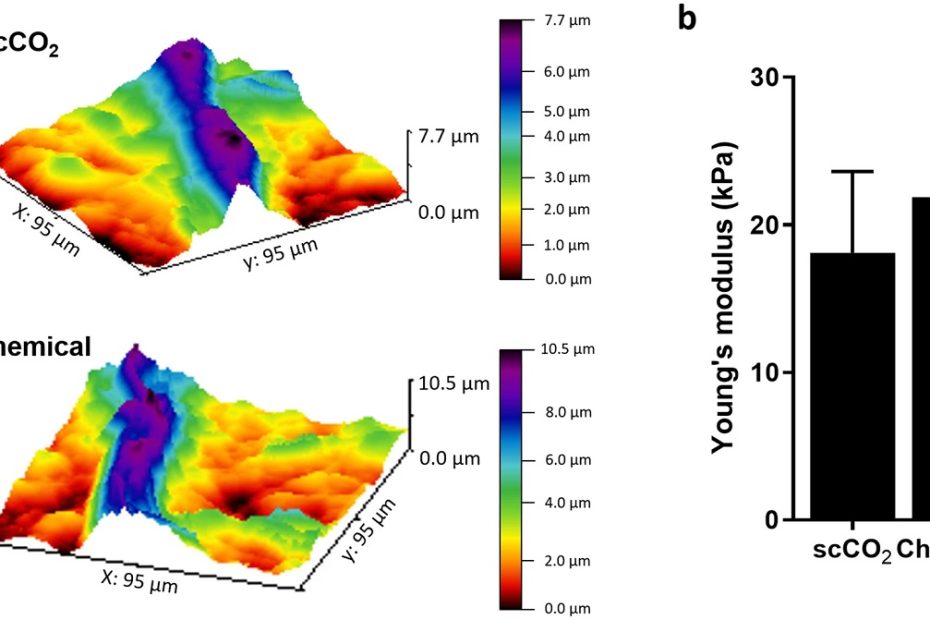
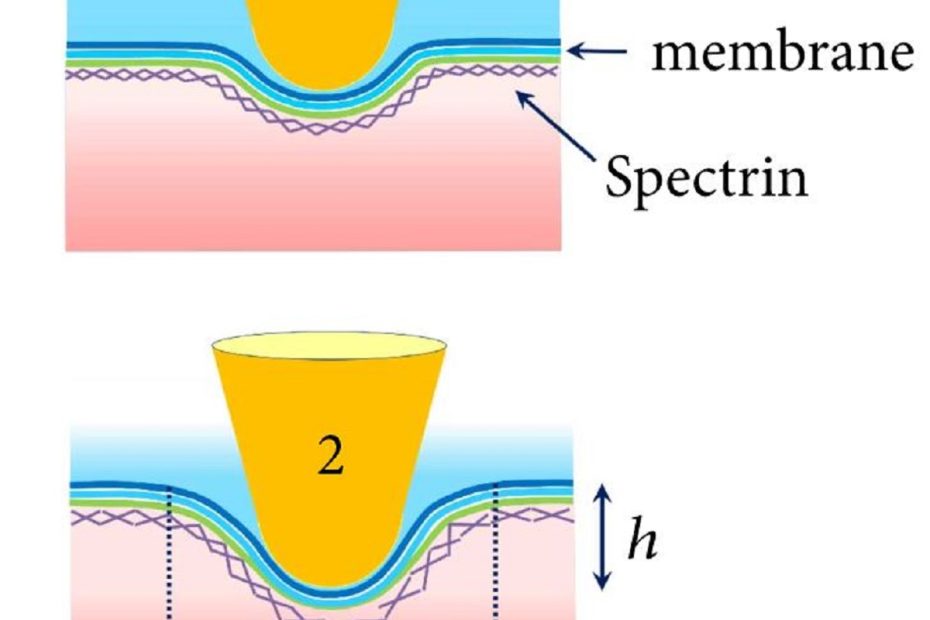
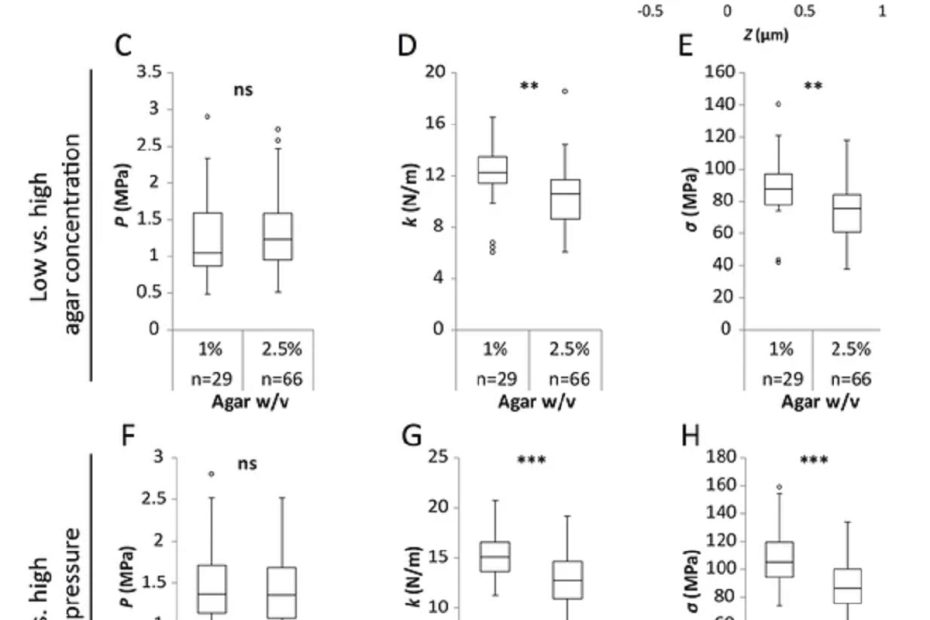
The interface between ice and alcohols analyzed by atomic force microscopy
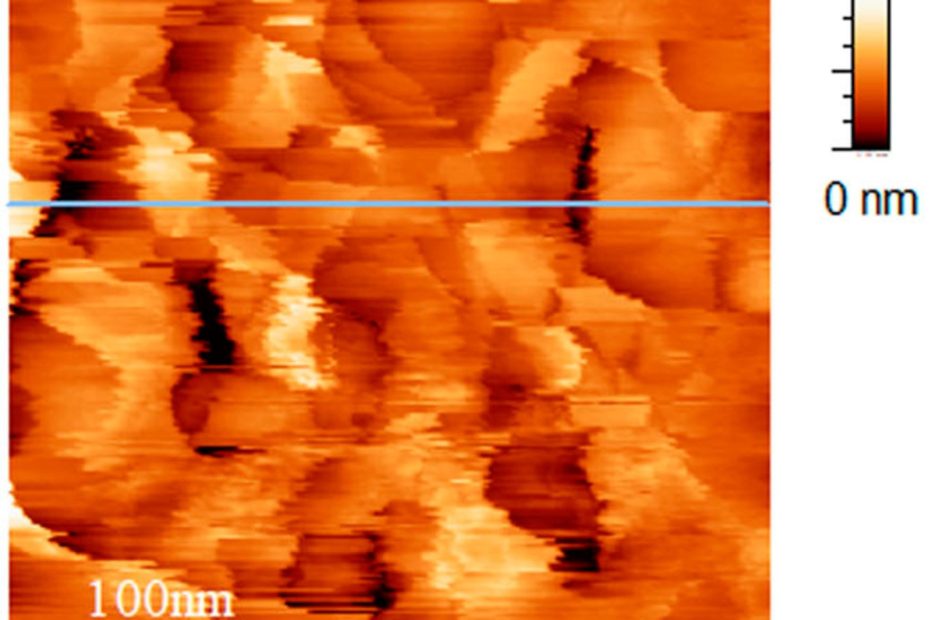
Ice plays a crucial role in our environment, with natural ice formations, such as glaciers, permafrost, river ice, and snow, strongly influencing the temperature, humidity,… Read More »The interface between ice and alcohols analyzed by atomic force microscopy